A Braun-Blanquet-féle módszer ellen felhozott legfontosabb érv a túlzott szubjektivitás. A klasszikus módszertannak az elsőtől az utolsó lépéséig olyan döntések meghozatala szükséges, amelyek jelentős mértékben függenek a cönológus személyes látásmódjától, korábbi tapasztalataitól, a vizsgálat körülményeitől. Ezek csökkentik a hagyományos cönológiai elemzések reprodukálhatóságát, az eredmények leellenőrizhetőségét, amelyek egy tudományos munka esetén elvárhatóak lennének.
 |
| A kvadrátok elhelyezése az egyik legfontosabb kérdés az vizsgálat során (http://lakesuperiorinterfaith.org/) |
A mintavételezett növényzet kijelölése
A Braun-Blanquet módszer preferenciális terepi mintavételt követ, ami azt jelenti, hogy a vizsgált területen a különböző jellegekkel rendelkező növényzeti foltok, állományok, nem azonos eséllyel kerülnek a mintába. Ebben az értelemben a minta nem reprezentatív, szigorú statisztikai értelemben vett becslésre és hipotézistesztelésre nem (vagy csak igen erős korlátokkal) használható fel. Ugyanakkor a klasszikus cönológiai szemlélettel vett minta gyakran átfogó képet ad egy-egy terület vegetációtípusairól, amennyiben a kvadrátok kihelyezését a terület vegetációtípusainak teljes dokumentálása irányította. Ebben az esetben a cönológiai minta nem sérti a statisztikai mintázatelemzés* feltételeit, noha az elemzés eredményeinek az értékelése ilyenkor is nagy körültekintést igényel.
*Megjegyzés: becslésről akkor beszélünk, amikor a minta alapján következtetünk a mintavételezett objektum (a statisztikai sokaság) valamely, egyetlen tulajdonságára. Egy példa: kérdésünk, hogy egy réten 1 négyzetméteren hány faj fordul elő átlagosan. A mintavételezett objektum (statisztikai sokaság) itt a rét, még pontosabban a réten potenciálisan elhelyezhető összes 1 nm-es négyzet. Nyilvánvaló, hogy nem tudunk felmérni minden lehetséges négyzetet, hiszen azok száma végtelen, ezért véletlenszerűen kihelyezünk elegendő mennyiségű négyzetet a réten, ezekben megszámoljuk a fajokat, majd ebből a mintából (melynek elemei az egyes felmért négyzetek) számoljuk ki az átlagos fajszámot. Ha a kvadrátok kihelyezése során nem sérültek a reprezentatív mintavétel feltételei, akkor joggal feltételezhetjük, hogy a mintából számolt átlagos fajszám közel áll a valósághoz. Ha volt egy előzetes várakozásunk az átlagos fajszámra nézve, akkor különböző módszerekkel tesztelhetjük azt a hipotézist, hogy a várakozásunk teljesül. Mintázatelemzésről akkor beszélünk, amikor a statisztikai sokaság sokféleségéről gyűjtünk információt. Például hogy a vizsgált réten milyen típusok különíthetők el a fajösszetétel alapján. Ilyenkor a mintavétel során a rét változatosságának minél teljesebb reprezentálása lehet a célunk, nem az "átlag" keresése.
A klasszikus cönológiai adatok statisztikai reprezentativitásáról szól a Folia Geobotanica folyóirat 2007 júniusi száma, de ezen kívül is számos cikk született és születik arról, hogy a mintavételi helyek kiválasztása miképp befolyásolhatja a vegetációelemzések eredményét. Mihalcová és mtsai (2011) cikkét ajánlom ebben a témában.
Amikor magunk végezzük a felvételezést, nyilván mi döntünk arról, hogy mit fog reprezentálni a minta. A felvételek preferenciális kihelyezése helyett elvileg többféle mintavételi elrendezésre (pl. véletlenszerű, rétegzett random, szisztematikus) van lehetőség, ezek tényleges megvalósíthatósága viszont gyakran rendkívül nehéz, és nem is feltétlenül célravezető. A cönológusok többsége megegyezik abban, hogy valamilyen releváns módon rétegzett mintavételt érdemes folytatni. Ezt azt jelenti, hogy egy előzetes osztályozást (sztratifikációt, "rétegzést") végzünk a vizsgált növényzeten a fő vegetációs mintázatok szerint, majd minden rétegben ugyanannyi felvételt készítünk. A rétegen belül nem a "tipikus" vagy "ideális" állományokat keressük, hanem próbálunk egy teljes, átfogó lefedettséget reprezentálni az egyértelműen atipikus, speciális, nem ismétlődő jellegű foltok elkerülésével. Ez egy enyhébb, megengedőbb preferencia, mintha konkrétan a tipikus foltokat keresnénk. Ugyanakkor ez sem mentes a felvételező szubjektumától: a rétegek definiálása, a felvételek pontos helyének megválasztása még mindig az egyéni döntésekhez kötött. A rétegeken belül véletlenszerű mintavétel is lehetséges, ez azonban nagyon nagy munkaigényű, kis földrajzi kiterjedésű vizsgálatok esetén valósítható csak meg.
A probléma régi felvételek hasznosításakor is felmerül. Például ha több országra kiterjedő elemzést szeretnénk végezni, de nincs módunk új, terepi mintavételre, akkor különböző adatbázisokból, vagy szakirodalmakból hozzájuthatunk azokhoz a felvételekhez, amelyeket az érintett területen más kutatók készítettek. Kérdés, hogy mennyire bízhatunk meg a felvételekben. A rendelkezésre álló felvételek köre ugyanis rendszerint egészen szövevényes úton áll össze. Szerepelhetnek benne a XX. század eleji "nagy öregek" első felvételei, akik a hazájuk vegetációjának (mai fogalmaink szerint) erősen szubjektív leírására törekedtek, szerepelhetnek benne egyes vegetációtípusokra koncentráló, szintetikus tanulmányokhoz készült felvételek, szerepelhetnek egy terület időbeli változásához készült megismételt felvételek, sőt, egy lelkes egyetemista szívének kedves területén készített felvételei is, amelyekre a diplomadolgozata épült - jó esetben. Általánosságban elmondható, hogy a könnyen megközelíthető vagy valami miatt közkedvelt tájakon, valamint az átlagos terepbotanikusok számára vonzó (természetes, specialista, valami miatt unikális...) vegetációtípusokban több felvétel készült, mint máshol. Nyilvánvaló, hogy egy ilyen minta nem ad reális képet egy nagyobb régió növényzetéről, mivel a területek és a típusok nem arányos mértékben jelennek meg benne. Azonban ha elegendő felvételünk van (ami egy ideális eset), akkor bízhatunk abban, hogy, ha nem is arányos mértékben, de minden terület és típus jelen van az adatbázisban. Ilyenkor a dolgunk csak annyi, hogy a túlreprezentált területekről és vegetációtípusokból származó felvételeket "megritkítsuk". Pl. ha egy területről van 100 felvétel, egy másikról csak 10, akkor a 100-ból kihagyunk 90-et, így mindkét helyről egyaránt 10 felvételünk lesz. Erről szól tulajdonképpen a rétegzett újramintavételezés (Knollová és mtsai 2005). A mintánkat felvételcsoportokra ("rétegekre") bontjuk annak megfelelően, hogy milyen változóra nézve látjuk egyenetlennek a reprezentativitását, majd a csoportokból azonos számú felvételt tartunk meg a tényleges elemzéshez. Rétegek definiálása és a megtartandó felvételek kiválasztása egy újabb feladat. A felvételek teljes változatossági tartományát egyenletesen lefedő újramintavételezés érhető el a HCR módszerrel (Lengyel és mtsai 2011).
Hasonlóan fontos lehet a felvétel készítésének időpontja. Bizonyos vegetációtípusokban, például zárt erdőkben ajánlott legalább évente kétszer feljegyezni a jelen lévő fajokat és tömegességüket. Ez azonban csak új felvételek készítésekor teljesíthető, régi adatokról gyakran nem derül ki, hogy hányszor látogatta meg a felvételező a kvadrátot. A felvételek készítésének pontos ideje gyakran nem is nyilvánvaló irodalmi adatok esetén. A legtöbb elemzésben az ebből fakadó potenciális torzítástól eltekintenek, pedig újabb kutatások szerint nagyobb körültekintéssel kell eljárnunk (Vymazalová és mtsai 2014).
Kvadrátméret
Nincs egyértelmű útmutatás a felvételek területére vonatkozóan sem. A hagyomány szerint az optimális kvadártméret ("minimal area") megállapításának módja, hogy térben egymásba ágyazott, különböző méretű kvadrátokban feljegyezzük a fajszámot, amely adatokból a fajszám-terület görbe ellaposodásának helye fogja megadni a keresett térléptéket. Ez a logika azonban egyértelműen téves, hiszen a fajszám a terület függvényében monoton nő, és megfelelő adattranszformációval lineárissá alakítható (Dengler 2009). Ma a cönológusok döntő többsége megegyezik abban, hogy ideális kvadrátméret nem létezik, csupán megegyezés kérdése, hogy milyen térbeli felbontásban vizsgálódunk. A gyakorlatban a legfontosabb, hogy tágabban értelmezett vegetációtípusokban (erdő, cserjés, rét, nyílt gyep...) következetesen azonos méretű kvadrátokat vegyünk fel. Az egyes szárazföldi vegetációtípusokban hagyományosan használt, és emiatt a jövőre nézve is ajánlott kvadrátméretek több cönológiai kézikönyv alapján a következők:
- nyílt gyepek: 4-25 nm
- zárt gyepek, kaszálórétek, magassásosok: 16-50 nm
- cserjések: 50-200 nm
- erdők: 100-400 nm
 |
| Az "optimális kvadrátméret" megállapításának hagyományos módja (www.scielo.br) |
Érdemes megnézni, hogy a cseh vegetációs adatbázisban milyen kvadrátméretekkel készült felvételek vannak vegetációtípusonként: Chytrý és Otýpková (2003) cikke.
Becslési skálák
A cönológusok által Európa-szerte használt becslési skálák többségében ordinálisak, vagyis a skála fokozatai közti különbségek számszerű értékkel nem bírnak, csak a köztük lévő relációnak (kisebb vagy nagyobb) van értelme. Hogy egy adott faj a skála melyik fokára kerül, az attól függ, hogy a felvétel területének mekkora részét borítja. Vagyis a felvételező a faj valódi borításértékét becsli meg fejben (a kvadrát területének százalékában), majd ez "butítja le" egy durvább, ordinális skálára. Például, ha egy faj a kvadrát területének 30%-át borítja, az a Braun-Blanquet-féle borítási kategóriák (ld. a korábbi bejegyzésben) közül a 3-as számmal jelzettnek felel meg, amelynek alsó határa 25%, felső határa 50%. A Braun-Blanquet-féle skála viszont ordinális természete miatt nem alkalmas számtani műveletekre, ellenben a valódi százalékos értékekkel. Bevett szokássá vált ezért az ordinális skálákon kódolt borítások átalakítása a kategóriák százalékos középértékére. Például a 25% és az 50% középértéke a 37.5%, ezért a 3-as borítású fajok 37.5%-ot kaphatnak az átalakítás után. A módszer hibája egyértelmű, hiszen az történik, hogy egy faj, amelynek tömegességéről a Braun-Blanquet kódok csak annyit árulnak el, hogy valahol 25 és 50% közötti területet borít be, kap egy 37.5%-ot, függetlenül attól, hogy a tényleges borításértéke mennyi volt. További inkonzisztencia forrása, hogy többfajta ordinális becslési skála is létezik. A Domin-skálán például egy 30%-os borítású faj a 6-os (25% és 33% között) kategóriába esik, míg egy 40%-os faj a 7-esbe (33%-50% között). A Domin-skálán a 6-os kategória középértéke 29%, a 7-esé 41.5%, ezeket a százalékokat kapná a két faj a százalékká transzformálás után. Ugyanakkor a Braun-Blanquet skálán mindkét faj a 3-as kategóriába esne és 37.5%-ot kapna. Az eltérő skálák használata és a többszörös transzformálás miatt ugyanaz a növényzeti állomány egymástól eltérő felvételekként is szerepelhetne egy közös elemzésben. A helyzetet tovább rontja, hogy a Braun-Blanquet-féle skálák egyes változatainál (köztük az eredetinél) nem csak a borításértéket, de a növény egyedszámát is figyelembe kell venni - ez a növényfajok tényleges borításának pontos reprodukálását végképp lehetetlenné teszi. Ezen a linken látható egy táblázat, amely összehasonlít néhány ordinális borítási skálát, illetve itt is egy:
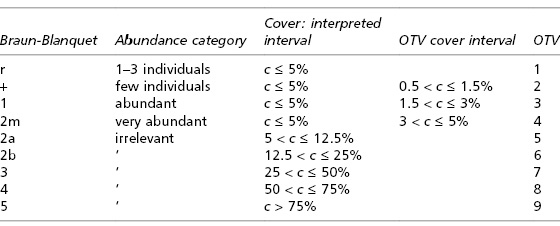 |
| A Braun-Blanquet-skála egy változatának és a van der Maarel-féle ordinális skála összehasonlítása (van der Maarel & Franklin: Vegetation Ecology, 2. kiadás, 2012) |
A modern cönológiai munkákban a fenti hiányosságokon többféle módon próbálnak úrrá lenni, több-kevesebb sikerrel. Az új adatok gyűjtése rendszerint már százalékos skálán történik, így az oda-vissza skálakonverzió problémájától mentesülünk. A régi, ordinális skálákon felvett adatokon azonban ez nem segít, marad a borításkategóriák középértékére való átalakítás, valamint az adatok utólagos ellenőrzése, mely során az elemzésből kihagyhatóak azok a felvételek, amelyeknél a hibás skálaalkalmazás gyanúja felmerül (pl. kiugróan alacsony vagy kiugróan magas borításértékek a transzformáció után). Az ordinális abundanciaadatok százalékokká alakítása védelmében megemlítendő, hogy az ordinális skálák többsége a nagyon alacsony értékeknél finomabb felbontású, mint a nagyobbaknál. Például a Braun-Blanquet skála 1-es kategóriája 1 és 5% közti borításért jár, vagyis 4% a kategória terjedelme. Ezzel szemben a 4-es fokozat 50 és 75% között található, tehát 25% terjedelmű. Ez összhangban van azzal a terepi tapasztalattal, hogy a 0%-hoz és a 100%-hoz közeli borítások becslése könnyebb, mint a közepes borításoké. A közepes borítású fajok százalékra pontos becslése a hamis pontosság látszatát kelti, mivel egy 45%-os borítású faj esetén 5% tévedés a legprecízebb terepi felvételezők esetén is előfordul, míg egy 5%-os borítású fajnál ugyanekkora hibát már jóval kevesebben követnek el. És ez nem csak a felvételezők tulajdonsága, általában ökológiai szempontból is lényegesebb egy ritka faj abundanciájának (abszolút értelemben) kis mértékű változása, mintha ugyanekkora eltérést találnánk a domináns faj borításában. A Braun-Blanquet skála tehát alkalmazkodik a tévedés várható mértékéhez és jelentőségéhez.
Elemző módszerek
Ezalatt az összes olyan statisztikai műveletet értem, amely az adatok feltárását szolgálja. A kezdeti, klasszikus cönológiai munkákban a felvételek és tabellák csoportosítása, a társulások elkülönítése a kutató terepi tapasztalatai alapján történt, ezek mellett csak alapszintű kvantitatív jellemzők játszottak szerepet (a fajok konstanciái és borításértékei). A modern cönológiai munkákban ezen lépések átláthatóbbá, reprodukálhatóbbá, "objektívebbé" tételére számos statisztikai módszert vezettek be. A felvételcsoportok elkülönítéséhez ma már nélkülözhetetlen valamilyen többváltozós osztályozó eljárás. Ezek a módszerek a felvételeket egymáshoz való hasonlóságuk alapján csoportosítják. A hasonlóság kifejezésére olyan indexek szolgálnak, amelyekkel páronként hasonlíthatóak össze a felvételek fajkompozíciójuk és a fajaik tömegességei alapján. A páronként összevetett felvételekből pedig többféle osztályozó algoritmus segítségével tárhatjuk fel az adatsorunkban rejlő csoportstruktúrát. Az osztályozás mellett a mintázat pontosabb megértését nagyban segítik az ordinációs módszerek, amelyek a csoportok helyett a fokozatos átmenetek, trendszerű mintázatok feltárására alkalmasak. Mára a hasonlósági indexek, az osztályozó és az ordinációs módszerek széles palettája áll a cönológusok rendelkezésére, az erről szóló irodalom is bőséges. Többé-kevésbé minden módszernek megvan a maga erőssége és gyengéje, és bár kétségtelen, hogy egyes eszközök értelmezhetőség szempontjából tetszetősebb eredményeket adnak, egyik módszer sem emelhető ki mint az egyedül üdvözítő, legalkalmasabb megoldás a cönológiai adatok elemzésére. Kimondottan nagy különbségek is lehetnek olyan alternatív módszerek alkalmazásakor, amelyek használata elméletben egyformán indokolható. Érdekes módon tehát a cönológus szubjektív döntése még mindig szükséges, csak nem konkrétan a felvételek csoportosítása során, hanem a csoportosításra használt módszer kiválasztásakor. A elemző módszerek mélyebb ismerete azonban nagy mértékben segít az általuk kapott eredmények magyarázatában, értelmezésében.
A csoportok leírása
A társulások domináns, konstans és diagnosztikus fajokkal való leírása tulajdonképpen nem sokat változott. Az egyetlen pont, ahol a szubjektivitásnak nagy szerepe volt, az a diagnosztikus fajok megítélése, hiszen azt kellett eldönteni, hogy két társulás összehasonlításakor egy fajnak mennyivel kellett gyakoribbnak lennie az egyikben, mint a másikban ahhoz, hogy az előbbire nézve diagnosztikus erőt tulajdonítsunk neki. A "diagnosztikus erő", amely tulajdonképpen a fidelitás (vagy hűség), mérőszámait a klasszikus cönológiai adatok elemzésében Botta-Dukát és Borhidi (1999), illetve Chytrý és mtsai (2002) vezették be. Ezek az indexek megmutatják, hogy egy adott faj előfordulásai mennyire erősen koncentrálódnak az egyes felvételcsoportokra. A rendelkezésre álló módszerek sokfélesége itt is nagy, ezek azonban többnyire hasonló eredményre vezetnek.
A fenti összefoglalóban csak vázlatosan próbáltam szemléltetni, milyen gyengéi lehetnek a Braun-Blanquet-féle társulástani munkáknak, és hogy milyen törekvések történtek és történnek ezek ellensúlyozására. A fentiekhez hasonló elméleti hiányosságok és a potenciális megoldások, a mintavételi és elemzési módszerek részletes ismertetése könyvnyi terjedelműre nyúlna, erre most nem vállalkozom, de remélhetőleg mások hamarosan megteszik helyettem. A biológiai, köztük cönológiai adatok elemzésekor számításba vehető statisztikai eljárások tárházáról magyarul olvasható egy kiváló könyv Podani János tollából, íme a link hozzá. A jelentős módszertani újítások, viták talán legfontosabb nemzetközi fóruma a Journal of Vegetation Science folyóirat. Korszerű, magas színvonalú elemzésekről szoktak megjelenni még cikkek a Folia Geobotanica, a Preslia, az Applied Vegetation Science és a Phytocoenologia lapokban. A hazai kutató műhelyek közül a MTA Ökológiai és Botanikai Intézetében (Vácrátót), az ELTE Növényrendszertani, Ökológiai és Elméleti Biológiai Tanszékén (Budapest) és a Pécsi Tudományegyetem Ökológiai Tanszékén folynak módszertani fejlesztésekkel kapcsolatos munkák, illetve modern módszereket alkalmazó társulástani kutatások. Remélem, hogy a leírtak alapján kitűnik, hogy a klasszikus cönológia közel évszázados, kezdeti gyakorlatához képest jelentős előrelépés történt és történik a tudományág elméleti, és ezzel összekapcsolódóan módszertani alapjainak megszilárdítása felé.