Akik magyar nyelven próbálnak korszerű, természettudományos ismereteket közzétenni, könnyen találkozhatnak azzal a nehézséggel, hogy egy idegen, rendszerint angol nyelvű, nemzetközi szakirodalomban közismert szakkifejezés nehezen ültethető át magyar nyelvre a kifejezés jelentéstartalmának, frappánsságának és használhatóságának megőrzésével, bármennyire egyszerű jelentéssel bírjon is. Éppen ez a helyzet a 'distance decay' nevű fogalommal.
A blogom nyilvánosságát megragadva javaslatokat várok arra, hogy milyen magyar nevet lehetne adni ennek a jelenségnek. Az angolban 'distance' = távolság, 'decay' =hanyatlás, rothadás. A javaslatokat hozzászólásként vagy a jobb oldali menüben a kapcsolatfelvételi űrlapon várom.
A '
distance decay' (mostantól DD-nek rövidítem - ha már nem magyar a szó, legalább legyen rövid) egy földrajzi kifejezés, amely egy teljesen triviális megfigyelésben gyökerezik, amit
Tobler (1970) után a
geográfia első törvényének is hívnak: "minden mindennel kapcsolatban áll, de az egymáshoz közeli dolgok erősebben, mint a távoliak". Ez tulajdonképpen a
térbeli autokorreláció definíciója. Nem tudom, van-e olyan földrajzi térbeli mintázat, amire ne lehetne találni olyan térbeli és időbeli vizsgálati léptéket, amelyben a fenti kijelentés teljesül - szerintem nincs. A jelenséget a fizikában tulajdonképpen az
általános tömegvonzás törvénye írta le elsőként, s a gravitációs modellek más tudományterületeken is analógiaként szolgáltak a távolság függvényében gyengülő interakciók leírására. Éppen így definiálhatnánk a DD-t:
a növekvő távolsággal gyengülő interakció közti függvénykapcsolat. A DD jelenséget általánossága miatt az élet legkülönbözőbb területein sikerült felismerni. DD-t mutat például
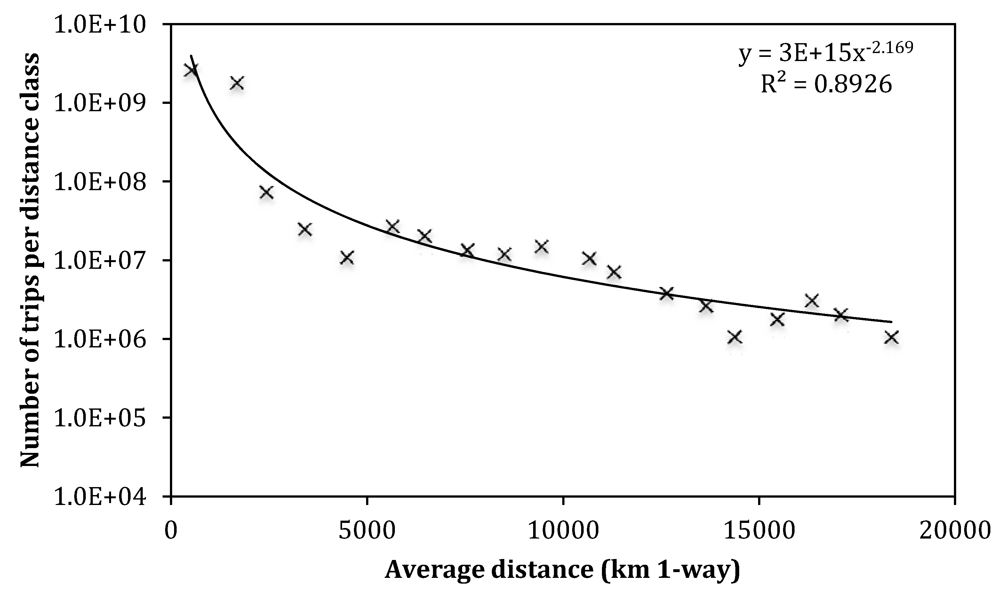
az utazások gyakorisága az út hosszának (vagyis az úticél távolságának) függvényében,
a koraszülöttek életben maradási valószínűsége a legközelebbi egészségügyi ellátóhelytől való távolság függvényében és a
kereskedelmi kapcsolatok erőssége a földrajzi távolság függvényében.
 |
Utazások száma adott távolságú célpontra
forrás: Peeters & Landré, Sustainability 2012, http://www.mdpi.com/2071-1050/4/1/42/htm |
 |
Különböző bolttípusok vásárlóinak száma a lakóhelyük távolsága szerint
forrás: http://lewishistoricalsociety.com/wiki2011/tiki-read_article.php?articleId=9 |
A térbeli autokorreláció ökológiai mintázatokban játszott fontos szerepét a XX. század utolsó évtizedeiben fogalmazták meg (
Legendre & Fortin 1989,
Legendre 1993), noha implicit módon már jóval korábban felismerték (pl.
MacArthur & Wilson 1967). A közösségi ökológia terén pedig
Nekola és White (1999) cikkéhez köthetjük a DD leírását, amely szerint a közösségek (most: adott területen előforduló fajok összessége) közti hasonlóság csökken a köztük lévő távolság növekedésével.
 |
Egy jellemző distance decay ábra a közösségi ökológia területéről: az x-tengelyen a közösségek földrajzi távolságai, az y-tengelyen a közösségek hasonlóságai, a pontok az összehasonlított közösségpárokat jelölik
forrás: Dexter et al., PNAS 2012, http://www.pnas.org/content/109/20/7787/F1.expansion.html |
A tapasztalat általában megerősíti azt a megfigyelést, hogy egy adott közösségtől minél messzebbre megyünk, átlagosan annál kevésbé várható, hogy hasonló fajösszetételű közösséget találjunk. Az azonban nagyon gyakran nem egyértelmű, hogy a hasonlóság csökkenésének mi az oka, illetve a különböző lehetséges okok milyen mértében járulnak ehhez hozzá. Amit tudunk, hogy a közösségek térbeli mintázata levezethető az egyes fajokéból (
Morlon et al. 2008,
Nekola & McGill 2014).
DD jelenség magyarázatához két modell lehet segítségünkre: a (1)
niche-modell és a (2)
korlátozott terjedési modell. A niche-modellben azt feltételezzük, hogy a vizsgált tájat különböző adottságú foltok alkotják, a foltok valamilyen léptékben térben aggregáltak (vagyis a környezetnek van térbeli autokorrelációja), a fajok élőhelyigényükben különböznek, valamint bármely nekik megfelelő élőhelyfoltban előfordulhatnak, terjedésüket egyedül az élőhely alkalmatlan mivolta korlátozhatja. Ebben az esetben tehát a közösség DD-je egy "külső" tényező, a környezeti DD lenyomata. A korlátozott terjedési modellben ezzel szemben azt feltételezzük, hogy a táj homogén a fajok megtelepedése szempontjából, a fajok élőhelyigénye nem különbözik, viszont terjedésük időben és térben korlátozott: új megtelepedések a régi megtelepedések közelében várhatóak, a teljes területen való elterjedéshez pedig hosszú időre van szükség. Eszerint a DD egy "belső" tényezőnek, a fajok korlátozott terjedőképességének köszönhető.
A valóságban rendszerint mindkét mechanizmus, tehát a niche-folyamatok és a korlátozott terjedés is felelős a tapasztalt összefüggés kialakulásáért. A DD jelenség tehát az alábbi tényezőkből vezethető le:
- a vizsgált területen a környezet heterogenitása, vagyis a területen az élőhelyfoltok különbözőségének mértéke: ez szabja meg, hogy az egyes fajok mennyi megtelepedésre alkalmas foltot találnak a területen, hiszen ha egy területen nagyon különböző élőhelyfoltok vannak, akkor azokon nagyon különböző élőhelyigényű fajok tudnak előfordulni, viszont minden élőhelyen talán egyik faj sem fordul elő.
- a vizsgált terület tájstruktúrája, vagyis az élőhelyfoltok térbeli mintázata: ezt fogja tükrözni a közösségek térbeli mintázata, amennyiben a terjedés azt lehetővé teszi.
- a fajok niche-szélessége, vagyis az, hogy mennyire ragaszkodnak egy bizonyos élőhelytípushoz (mennyire generalisták vagy specialisták): a generalista fajok többféle élőhelyen is előfordulnak, így a térbeli mintázatukat nem annyira az élőhelyfoltok térbeli struktúrája, hanem a terjedési potenciáljuk határozza meg. A specialista fajok egy adott élőhelytípushoz ragaszkodnak, és jó terjedőképesség esetén pontosan követik annak foltosságát.
- a fajok terjedőképessége: minél jobban terjed egy faj, annál inkább a megfelelő élőhelyek térbeli mintázatát követi, míg a rosszul terjedő fajok nem jutnak el minden potenciálisan megfelelő foltba, vagy nem maradnak fenn ott hosszú távon. (Pl. egy hím madárnak nem elég eljutni egy élőhelyre a faj tartós megtelepedéséhez, mert ehhez kell egy tojó is...)
A DD-re ható tényezőkkel Nekola & White (1999) és Soininen et al. (2007) is foglalkoznak, mindkét cikkben megfogalmaznak általánosságokat arról, hogy mely tényezők eredményeznek "meredek", "erős", csökkenést a hasonlóságban, vagy nagyfokú térbeli strukturáltságot. Később bemutatom, hogy a DD milyen sokféle mérőszámmal írható le, amelyek mind a csökkenés mértékének kifejezésére szolgálnak, de valójában nem egészen ugyanazt fejezik ki, és nem is függetlenek egymástól. Emiatt igyekszem óvatosan fogalmazni azzal kapcsolatban, hogy egyes tényezők hogyan hatnak a várható DD-re.
 |
| A változatos domborzatú, magashegységi tájban a közösségek hasonlósága kis távolságon belül is hirtelen lecsökkenhet |
A niche-modell reális magyarázatot nyújt a DD-re, ha:
- a környezeti (a fajok szempontjából) heterogén, vagyis a fajok specialisták,
- a környezeti háttérváltozók térben autokorreláltak,
- a fajok jól (csaknem korlátlanul) terjednek.
A korlátozott terjedési modell ad jó magyarázatot, ha:
- a környezet (a fajok szempontjából) homogén, vagyis a fajok generalisták,
- a háttérváltozók nem autokorreláltak (ez részben a homogenitásból is fakadhat),
- a fajok rosszul terjednek.
Nincs jelentős DD, ha:
- a környezet (a fajok szempontjából) homogén vagy heterogén, de akkor a változók a vizsgált léptékben nem autokorreláltak,
- a fajok jól terjednek.
 |
| A kis holdruta (Botrychium lunaria) jól terjedő, de specialista faj |
Hogy konkrét esetekben a fenti szempontok hogyan teljesülnek, az az adott vizsgálat módszertani döntéseitől is függ, mindenek előtt a mintavételi egységek méretétől és a mintavételezett terület nagyságától. Ugyanaz a vizsgálati objektum teljesen más DD-t mutathat, ha eltérő térléptékben vizsgáljuk. Pl. ha Magyarország növényzetét vizsgáljuk, más térbeli mintázatok tűnnek fel, ha a mintavételi egységeink 1 m2-esek és ha 1 km2-esek. Általánosságban igaz, hogy minél nagyobb a mintavételi egység és a mintavételezett terület, annál valószínűbb, hogy erős DD összefüggést kapunk.
Ennyi elmélet után jogosan merül fel a kérdés:
mire jó ez az egész? A válasz(om): az, hogy valamilyen élő közösség, pl. egy terület növényzete, milyen térbeli változatossággal (
béta diverzitással) rendelkezik, alapvető kérdés a természetet leíró, ökológiai kutatásokban. Az erről szerzett ismeretek pedig fontosak lehetnek természetvédelmi értékek minősítése, veszélyek felmérése, területek védelme szempontjából. Például ha egy területről kimutattuk, hogy a rajta élő fajok terjedőképessége alacsony, akkor a természetvédelmi kezeléseknek kiemelt figyelmet kell fordítaniuk arra, hogy a fajok potenciális élőhelyei között ökológiai folyosókat tartsanak fenn, valamint fokozottan kell ügyelniük a lokális kihalások elkerülésére. A béta diverzitás térbeli elemzése szintén hozzásegít olyan alapvető biológiai folyamatok megértéséhez, mint a fajképződés, a klímaváltozások alkalmával történő flóra- és faunavándorlások és a biológiai inváziók. Hogy a gyakorlati célok számára is fontos információk birtokába technikailag hogyan juthatunk a DD modellek segítségével, egy későbbi bejegyzésben fogom leírni.